In the year 2000 the draft human genome sequence was announced by Tony Blair and Bill Clinton. It was said to be complete in 2003, in time for the 50th anniversary of the discovery of the structure of DNA. Well actually it wasn’t quite finished. Actually it’s still not finished. Besides the tweaking that still goes on here and there, there are still big gaps. And what’s in these gaps sometimes has a significant role in cancer.
The biggest gaps are centromeres. Centromeres and telomeres are made of what is known as repetitive DNA, and this is hard to sequence. There’s more detail below.
Cancer chromosomes often have centromere or telomere abnormalities. In fact these abnormalities can cause cancer or make it progress faster.
In cancer research there’s a big push to sequence the genomes of different types of cancer to try and understand the many different DNA changes that can cause cancer. Some researchers try to understand telomeres and centromeres and their role in cancer, but in cancer sequencing projects, and also in diagnostics, centromeres and telomeres are pretty much ignored. Although they’re difficult to sequence, the repetitive DNA does make them easy to study by some other techniques.
One of the goals of personalised medicine is to be able to read a person’s complete genome. For cancer this would include the abnormal cancer genome. But at the moment these gaps mean that we can’t describe the abnormal cancer chromosomes from end to end by sequencing them. The approach I use allows me to work out what’s in each chromosome and discover telomere and centromere abnormalities.
HOW DOES MY RESEARCH FIT IN?
By looking at things that most people don’t worry about I’ve overturned a few assumptions and made some unexpected discoveries, particularly about centromeres in leukaemia.
A normal chromosome has one centromere. I found that chromosomes with two centromeres are more common in acute myeloid leukaemia (AML) and myelodysplastic syndromes (MDS) than was thought. I found that most of these chromosomes with two centromeres were probably made by two chromosomes joining together.
Telomeres are at the ends of chromosomes and stop them from sticking to each other, so when the telomeres are eroded, chromosomes can join together. They can be eroded by exposure to chemical toxins, cancer drugs and radiation. So it’s interesting that the leukaemias that are caused by these exposures have more of these two-centromered chromosomes than other leukaemias.
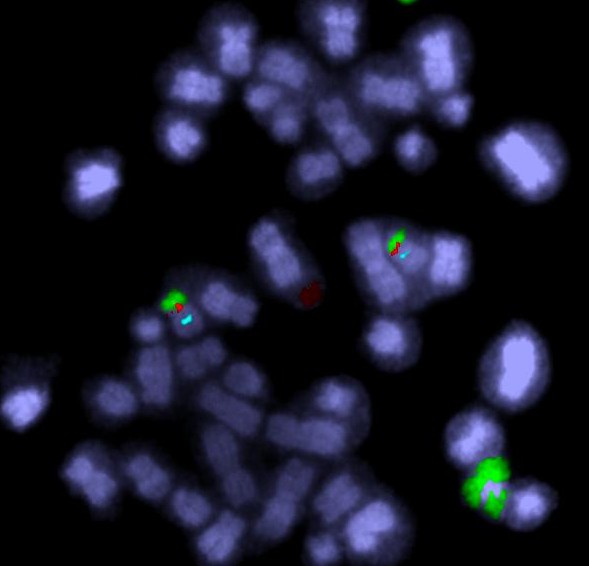
Fluorescent DNA probes can label up centromeres (blue) and genes (red). In this image there is also a chromosome 20 paint – the green regions are from chromosome 20.
MORE DETAIL ON THE SCIENCE
Telomeres and centromeres are made of highly repetitive DNA and make up some of the gaps in the human genome sequence.
Sequencing a genome is like reading a story. But first we cut the book up into tiny fragments. We read them piece by piece, then try to join the pieces together to make the story, by matching the overlapping parts. Where this approach falls apart is that some sections are repetitive. Some pages are made up of a single word or phrase repeated over and over and over and over and over and over (I won’t repeat that hundreds of thousands of times, but you should get the picture). So if a lot of fragments just say the same thing “over and over and over”, it’s very hard to put them together meaningfully.
The centromere guides the chromosome to the two daughter cells during cell division. A normal chromosome has one centromere. When a chromosome has two centromeres (we call this a dicentric chromosome), the chromosome can be pulled in opposite directions, breaking the chromosome and causing more chromosome disorganisation.Telomeres cap the ends of normal healthy chromosomes. One of their functions is to stop the chromosomes from sticking to each other. So when the telomeres are lost or eroded the chromosomes can join together. That’s one way of creating a chromosome with two centromeres.
Telomere loss is a natural part of ageing. There are also many environmental and lifestyle factors that are thought to affect telomere length. Short telomeres are thought to be a cancer risk because dicentric chromosomes are more likely to arise.
Telomeres and centromeres are very important parts of a normal chromosome. You could say they hold the chromosome together. They have a lot of influence on whether chromosomes are normal and stay normal.
FURTHER READING
Murnane JP 2012. Telomere dysfunction and chromosome instability. Mutat Res. 2012 Feb 1;730(1-2):28-36. (Open access)
MacKinnon RN and Campbell, LJ. 2011. The role of dicentric chromosome formation and secondary centromere deletion in the evolution of myeloid malignancy. Genetics Research International. Article ID 643628. (Open access)
MacKinnon RN, Duivenvoorden HM and Campbell LJ. 2011. Unbalanced translocation of 20q in AML and MDS often involves interstitial rather than terminal deletion of 20q. Cancer Genet. 204(3):153-61.


